| Главная » Статьи » ОТРЯД КИТООБРАЗНЫХ » Подотряд зубатых китов |
ОТРЯД КИТООБРАЗНЫХ ORDO CETACEA BRISSON, 1762
| К отряду принадлежат высокоспециализированные млекопитающие, приспособившиеся к постоянной жизни в воде. Размеры средние и крупные (наибольшие среди млекопитающих). Длина тела животных от 1.1 м. масса 30 кг у некоторых дельфинов, до 33 м. и 150 т. у голубого кита 1. Наименьшее сопротивление воды при плавании обеспечивается хорошо обтекаемым телом, торпедообразным или каплевидным, вытянутым или несколько укороченным (рис. I). Этому способствуют также редукция ушных раковин и расположение сосков в особых карманах и пениса в специальной сумке. Передние конечности превратились в ласты (грудные плавники), которые служат в основном рулями. Свободные Задние конечности подверглись атрофии. На конце латерально уплощенного хвостового стебля имеются парные, горизонтально расположенные бесскелетные хвостовые лопасти. Хвостовой отдел туловища служит основным локомоторным органом. У большинства видов на спине находится непарный бесскелетный спинной плавник, который служит своего рода стабилизатором при плавании. Голова часто массивная, более или менее удлиненная. Ее терминальная часть кончается тупо, заострена или имеет вытянутый рострум — «клюв». Почти без видимого шейного перехвата голова переходит в туловище, постепенно без резкой границы сужающееся в хвостовой стебель. Характерно для китообразных, что у них нет сплошного волосяного покрова. У усатых китов на морде расположены отдельные щетннковидные волоски, обладающие типичным строением вибрисс наземных млекопитающих; по-видимому, они служат органом осязания и играют определенную роль при отыскании массовых скоплений планктона. Во взрослом состоянии у зубатых китов вибриссами обладают только Platanista gangetica и Inia goeffrensis, живущие в мутной речной воде; вибриссы у них наблюдаются не только на морде, но и на туловище. У большинства других зубатых китов (кроме белухи, нарвала) вибриссы имеют только эмбрионы. Кожные железы полностью отсутствуют, за исключением млечных желез. В коже на внутренней поверхности эпидермиса расположены многочисленные ячеи, и которые поднимаются дермальные сосочки. Некоторые ученые предполагают, что своеобразное строение кожи китов обусловливает возникновение ламинарного потока воды вокруг плывущего животного, что позволяет затрачивать минимум усилии при плавании и развивать большую скорость. Окраска тела обусловлена пигментом, локализующимся в основном в эпидермисе. У некоторых китообразных она подвержена возрастной и изменчивости (например, белуха, нарвал). Лопасти хвостового плавника и спинной плавник относятся к кожным образованиям. Наряду с обычными артериями и венами в плавниках проходят характерные только для них комплексные сосуды, каждый из которых состоит из крупной артерии с очень толстой средней оболочкой, окруженной 10-13 более мелкими тонкостенными венами. В основании спинного плавника дельфина-белобочки таких сосудов насчитывается свыше двух десятков, в хвостовом и грудном плавниках их значительно меньше (Томилин, 1957). 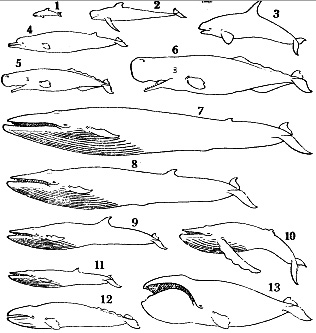 Рисунок 1. Относительные размеры и форма тела китообразных: 1 - дельфин-белобочка, 2 - гринда, 3 - косатка, 4 - северный плавун, 5 - кашалот самка, 6 - кашалот самец, 7 - голубой кит, 8 - финвал, 9 - сейвал, 10 - горбатый кит, 11 - малый полосатик, 12 - серый кит, 13 - гренландский кит В черепе китообразных кости располагаются симметрично или асимметрично. В последнем случае кости правой стороны расширены и смещены влево, а левой — утолщены. Одни кости могут надвигаться на другие. Мозговой отдел черепа укорочен и округлен. Верхнезатылочная кость сильно развита и спереди причленяется к лобным, в то время как теменные смещаются в стороны. Носовые кости невелики по размерам, так что наружные костные носовые отверстия сдвинуты далеко назад и располагаются непосредственно перед мозговым черепом. Носовые ходы короткие и проходят более или менее вертикально. Верхнечелюстные и межчелюстные кости и сошник сильно вытянуты и образуют рострум. Костное небо удлинено за счет крупных крыловидных костей, вследствие чего хоаны оказываются смещенными назад. Нижние челюсти длинные и имеют рудиментарный венечный отросток, которого иногда нет вовсе. 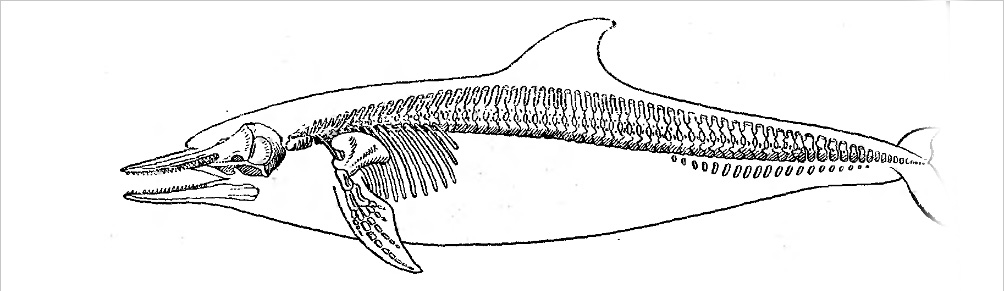 Рис. 2. Скелет и контуры дельфина-белобочки — Delphinus delphis Во взрослом состоянии зубы (гомодонтного типа) имеют только представители подотряда зубатых китов. У усатых китов закладки зубов имеются только у эмбрионов. Для взрослых усатых китов характерны особые роговые образования эпителия неба — так называемые «усы», функционально заменившие зубы. «Усы» служат для отцеживания мелких планктонных организмов, которыми питаются эти киты. Акт пережевывания пищи у китообразных утрачен. Кости посткраниального скелета в большинстве своем губчатого строения с большим содержанием жира. В поясе передних конечностей отсутствует ключица (рис. 2). Лопатка широкая, вееровидная со слабо развитым гребнем. Грудина сильно варьирует по форме и расчлененности. Плечевая кость чрезвычайно укорочена. Локтевая и лучевая кости также сильно укорочены, уплощены и расширены. В связи с тем что нагрузка на конечность происходит перпендикулярно к ней, конечность уплощается в том же направлении. Подвижное сочленение находится только в плечевом суставе. Все кости свободной конечности имеют общий кожный покров и сочленены между собой неподвижно, а иногда и срастаются друг с другом. Чаще, по сравнению с другими млекопитающими, наблюдается слияние карпальных костей. Пальцев 4 или 5. Средние обладают увеличенным количеством фаланг, крайние — уменьшенным. При сравнении числа фаланг пальцев кисти взрослых китообразных с эмбрионами отмечается их сокращение с возрастом. Закладка задних конечностей появляется на ранней эмбриональной стадии, но вскоре исчезает. У взрослых китообразных от пояса задних конечностей сохраняются только рудименты таза, расположенные в мускулатуре по большей части в виде двух костных стерженьков (слившихся подвздошной, седалищной и лобковой костей). Связь с позвоночником эти кости утрачивают, но к ним прикрепляются мышцы мочеполовой системы, в связи с чем, очевидно, и не происходит их полная редукция. Позвоночник состоит из четырех отделов: шейного, грудного, поясничного и хвостового. Крестцовые позвонки отсутствуют вследствие утраты задних конечностей. Шейный отдел представлен семью сильно укороченными позвонками, которые могут сливаться между собой в одну или несколько групп. В остальных отделах количество позвонков непостоянно. В хвостовом отделе развиваются гемальные дуги, аналогичные дугам низших позвоночных. У быстроплавающих китов число хвостовых позвонков возрастает. Число ребер варьирует от 10 до 17 пар. Нормальное сочленение с позвонками имеют передние 2—8-я пары; у задних ребер головка и шейка отсутствуют. К грудине присоединяются передние 1—8-я пары ребер. Для глубоконыряющих китов характерно уменьшенное число ребер, прикрепляющихся к грудине (у кашалота — три) или развитие в ребрах хрящевых отделов (клюворылы). Грудная клетка в отличие от большин¬ства наземных млекопитающих не сжата с боков, а округлая в поперечнике пли несколько уплощена дорзо-вентрально. Язык хорошо развит, мягкие губы отсутствуют. Слюнных желез нет или они рудиментарны. Желудок сложный, состоит из 3-5 отделов. Кишечник достигает различной длины: он может быть в 5-6 раз длиннее тела (усатые киты, клюворылы) или в 15-16 раз (кашалот, афалина) и до 32 раз (дель¬фин Ла-Платы). Печень относительно невелика, желчного пузыря нет. Поджелудочная железа удлиненная и слабо дольчатая, значительно реже разъединенная. Не протоки открываются в желчный проток. Наружные ноздри парные у усатых китов, непарные (одна ноздря) у зубатых. Они смещены на верх головы и обладают особыми клапана¬ми, запирающими эти отверстия при нырянии. Трахея и бронхи укорочены, что способствует убыстрению акта дыхания. Легкие однодольчатые с сильно развитой гладкой мускулатурой. Число альвеол относительно больше и размеры их намного крупнее, чем у наземных млекопитающих. Китообразные способны подолгу не дышать, находясь под водой от 2-10 до 30-40 мин (по некоторым данным, до 2 ч). Длительность ныряния у них обеспечивают значительная емкость лёгких, слабая чувствительность дыхательного центра к накоплению в организме углекислоты, повышенное содержание миоглобина. Кислородная емкость крови не¬сколько увеличена за счет более высокого содержания гемоглобина в крови и повышения его концентрации в эритроцитах. Процесс дыхания китообразных обычно можно подразделить на вы¬дох после длительного ныряния, промежуточные короткие вдохи и выдохи и глубокий вдох перед длительным нырянием. Во время промежуточных вдохов и выдохов кит ныряет неглубоко, плывет почти всегда по прямой линии, дышит через правильные промежутки времени. Число промежуточных вдохов-выдохов различно у разных видов и тем больше, чем дольше остается животное под водой во время основного ныряния. Выдох может начинаться у поверхности под водой, в результате чего выдыхаемый с силой воздух образует водяной фонтан, форма и раз¬меры которого в общем характерны для различных видов 2. Некоторые китообразные способны нырять на значительную глубину (кашалоты — свыше 1000 м). Быстрый подъем нырнувшего животного с большой глубины на поверхность воды должен вызвать выделение из крови газообразного азота, растворенного в ней вследствие повышенного давления при нырянии. Пузырьки азота могут закупоривать кровеносные сосуды, вызывая кессонную болезнь и даже смерть. Однако у китообразных кессонной болезни не бывает. Вероятно, это объясняется тем, что количество азота в легких не намного превышает азотную ем¬кость тканей кита, а воздух в процессе ныряния в легкие больше не по¬ступает (в противоположность тому, что имеет место у водолазов). Головной мозг китообразных во многих отношениях высоко дифференцирован. Однако но ряду показателей сохраняет весьма примитивные признаки, которые, видимо, были характерны для мозга млекопитающих исходного типа. Масса головного мозга в абсолютных цифрах (белуха — 1,6-2,3 кг; кашалот —7,8-9,2 кг) достигает наибольшей величины среди млекопитающих, но относительно массы всего тела она незначктельна (морская свинья — 0,8%, голубой кит —0,007%). В укороченном, имеющем округлую форму головном мозгу крупные извилины и бороз¬ды располагаются в продольном направлении. Характерно сильное развитие коры полушарий переднего мозга. Происходит редукция Rhinocephalon. Bulbus olfactorius у большинства китообразных отсутствует (Bulbus olfactorius описана у бутылконоса, голубого кита и финвала). Глаза небольшие. Роговица и склера достигают значительной тол¬щины. Хрусталик имеет своеобразную округлую форму. Веки неразвиты. Зрение китообразных, по-видимому, монокулярное, они близоруки. Слезные железы редуцированы, отсутствует и слезно-носовой проход. Жирный секрет гардеровой железы предохраняет глаз от механического и химического воздействия воды. Имеются конъюнктивные железы, не встречающиеся у других млекопитающих. Органы слуха сильно видоизменены. Ушная раковина редуцирована. Наружный слуховой проход открывается позади глаза маленьким отверстием. Интересно мнение, что рудиментарный слуховой проход может служить самостоятельным чувствующим органом, воспринимающим изменение давления. Барабанная перепонка выгнута наружу (усатые киты) или внутрь (зубатые киты). С внешней стороны барабанная перепонка усатых китов покрыта своеобразной ушной пробкой, состоящей из ороговевшего эпителия и ушной серы. Китообразные способны улавливать широкий диапазон звуковых волн от 150 до 120—140 тыс. Гц (Slijper, 1962), т. е. даже ультразвуковые колебания. Высокая степень развития слуховых отделов мозга зубатых китов свидетельствует об особой остроте их слуха, почти уникальной среди млекопитающих; у усатых китов слух хуже по сравнению с наземными млекопитающими. Китообразные способны к эхолокации, как и летучие мыши. В связи с тем что голосовые связки у китообразных отсутствуют, они не могут издавать звуки обычным для млекопитающих способом. Возможно, звуки производятся в результате вибрации нижней части перегородки между носовыми мешками или колебаниями складки наружного клапана в результате пропускания воздуха из дорзальных носовых мешков. Дельфины способны испускать серии коротких звуковых импульсов, длительность которых составляет 1 мс, а частота следования меняется от 1—2 до нескольких сот герц. В кровеносной системе китообразных сильно развита артериальная чудесная сеть (retia mirabilia), находящаяся на вентральной стороне позвоночного стебля, в области шеи, грудной клетки, между ребрами, в основании головного и вокруг спинного мозга. Она образована огромным числом извилистых артерий мускульного типа, связанных между собой многими анастомозами. Имеется к венозная чудесная сеть, рас¬полагающаяся в основании черепа и в области живота. Температура тела китообразных сходна с температурой наземных млекопитающих и колеблется в пределах от 35 до 40°С (верхняя граница отмечалась у раненых китов или пойманных после преследования дельфинов). Сохранение высокой температуры тела в воде, проводящей тепло во много раз лучше, чем воздух, осуществляется толстым слоем под¬кожной жировой клетчатки в коже. Почки китообразных много дольчатые, относительно более крупные, чем у наземных млекопитающих. Мочевой пузырь небольшой и в шейке не имеет сфинктера, что способствует частому мочеиспусканию небольших порций мочи, которая, возможно, может служить сигналом для других китов. Китообразные, по-видимому, не пьют морской воды. Они получают необходимую влагу из пищи, используя и оксидазную воду. Семенники располагаются в брюшной полости. Из дополнительных имеется только простата. Os penis отсутствует. Матка двурогая. У самок усатых китов половое и заднепроходное отверстия разделяются друг от друга значительным пространством, а у зубатых они расположены в едином углублении и окружены общим сфинктером. Самцы посто¬янно или очень подолгу в течение года способны к оплодотворению. Вы¬сказываются предположения, что овуляция у китообразных провоцируется половым актом. У самок в начале беременности в матке могут находиться два или три зародыша, из которых вскоре остается только один. Плацента диффузного типа. Роды происходят под водой. Детеныш рождается вполне развитый, способный к самостоятельному передвижению. Пропорции его тела весьма сходны с пропорциями тела взрослых китов, а размеры достигают 1/2—1/4 длины тела матери. Самки некоторых китообразных могут быть оплодотворены вскоре после родов во время лактационного периода. Кормление детенышей совершается под водой, длительность каждого приема нищи — несколько секунд. Молоко вбрызгивается в ротовую полость детеныша сокращением особых мышц самки. Млечные железы самки располагаются по бокам полового отверстия. Два соска (по одно¬му с каждой стороны) залегают в щелевидпых складках и только в период лактации выдаются наружу. Самки китов продуцируют различное количество молока в сутки: от 200—1200 г у дельфинов до 90—150 л у фннвала и 200 л у голубого кита (Слепцов, 1955). Молоко густое, обычно кремового цвета. Характерно, что его поверхностное натяжение в 30 раз больше, чем у воды, что особенно важно, так как струя молока не расплывается в воде. Питательность китового молока весьма высока. Рост детенышей во время вскармливания их молоком происходит быстро. Например, детеныш голубого кита за 7 месяцев жизни вырастает с 7 до 16 м, т. е. среднесуточный прирост в длину составляет 4,5 см. Половой диморфизм проявляется главным образом в различной дли¬не тела самцов и самок. Самки усатых китов крупнее самцов, а большинства зубатых, наоборот, меньше. Диплоидное число хромосом у 12 видов зубатых китов и 4 видов усатых (сейвала, малого полосатика, финвала и серого кита) равно 44, а у кашалотовых — 42. Распространены во всех океанах и в большинстве морей Земного шара. Факторы, определяющие размещение китообразных,— наличие пищи и температура воды. Некоторые виды распространены широко и встречаются как в теплых, так и в холодных морях (некоторые виды из семейства дельфинов), другие имеют меньший ареал (серые киты обитают в субтропических, умеренных и холодных водах северной половины Тихого океана и в Чукотском море), ареал третьих еще более ограничен (нарвал не покидает вод Арктики), наконец, ареал речных, озерных и эстуарных форм совсем незначителен. Большинство видов — стадные животные; держатся группами от не¬скольких голов до сотен и тысяч особей. Встречаются как возле побережий, так и в открытом море. Представители некоторых видов способны подниматься вверх по крупным рекам, впадающим в море, а отдельные виды постоянно живут в реках. У большинства китов питание специализировано, и среди них встречаются планктофаги, теутофаги, ихтиофаги и сакрофаги. Кормятся массовой или штучной добычей. Среди китообразных есть быстрые пловцы (косатки, многие дельфины) и относительно медленно передвигающиеся (серые киты). Большинство китов постоянно держатся в поверхностных водах. Некоторые, например кашалот, могут нырять на значительную глубину. Численность различных видов китообразных неодинакова. Многие из них весьма многочисленны и могут встречаться тысячными стадами (дельфин-белобочка), другие, напротив, очень редки и встречи с ними отмечены всего несколько раз (некоторые представители рода ремнезубов, карликовый кашалот, карликовая косатка). Естественные колебания численности не изучены; вряд ли они могут быть значительными. Чрезмерно интенсивный промысел пагубным образом сказывается на численности китов, заметно снижая ее, и в некоторых случаях может угрожать полным уничтожением этих животных. Так, ничтожная в настоящее время численность гренландских китов — следствие хищнического выбоя этих многочисленных когда-то животных. Для большинства видов характерны периодические миграции. У не¬которых видов протяженность миграционных путей относительно неве¬лика (азовочерноморская морская свинья — из Азовского моря в Черное и обратно); у других огромна (некоторые крупные киты — от тропических вод до высоких широт). Китообразные — преимущественно моногамы. Периоды спаривания и щенки обычно растянуты во времени. Рождают одного, редко двух детенышей. Сильно развит материнский инстинкт. Врагов, кроме человека и косатки, практически не имеют. Остатки дельфинов находили в желудках тигровых и гренландских акул. Почти 100% особей заражены эндо- и эктопаразитами. Эктопаразиты: китовые вши — Cyamus, копепода — Penella, усоногие ракообразные— Conchoderma — свойственны особенно крупным китам. Практическое значение в недалеком прошлом было довольно вели¬ко. Некоторых из китообразных и в настоящее время добывают ежегодно в больших количествах (кашалоты), других промышляют лишь случайно. Почти все органы кита используются для выработки ценных пищевых и технических продуктов. Промысел китообразных может вес¬тись только при строгом соблюдении мер, необходимых для поддержания их численности. В настоящее время в целях сохранения китообразных целесообразно прекратить их промысел на ряд лет. Ископаемых остатков китообразных известно относительно немного. Наиболее примитивные из известных нам китообразных (Protocetus, подотряд Archaeoceti) были найдены в среднем эоцене Северной Африки. Представители подотряда Archaeoceti образовали специализированную ветвь, дивергировавшую от ранних предковых форм и развивавшуюся самостоятельно до полного исчезновения в верхнем эоцене. Ископаемые остатки из позднего среднего и верхнего эоцена свидетельствуют, что Archaeoceti имели две основные группы, различающиеся по размерам и скелету. Представитель одной из них — Basilosaurus, имел змееподобное тело длиной 17- 22 м, специализированный позвоночник, способный изгибаться, как у змеи, и горизонтально расположенный маленький хвостовой плавник. Представители другой группы — Dorudon, Prozeuglodon, были значительно меньших размеров и походили на дельфинов. Их позвоночник напоминает позвоночник современных дельфинов. О предках китообразных нет единого мнения. Ряд ученых высказываются в пользу происхождения китообразных от копытных, с которыми они имеют ряд общих черт: диффузную плаценту, двурогую матку, сложный желудок. Анализы числа хромосом и изучение реакции преципитации белка крови китообразных подтвердили эту гипотезу. Другие исследователи на основании сходства в строении скелета и зубной системы древних китообразных с креодонтами предполагают, что китообразные произошли от последних. Высказывается предположение, что родоначальниками китообразных явились болee древние формы, чем креодонты, а именно меловые насекомоядные. Некоторые ученые (например, Beddard, 1900; Клей- пенберг, 1958: Андерсон и Джонс, 1967) предполагают дифилитическое происхождение отряда китообразных. Эволюция усатых и зубатых китов, их мнению, шла путем конвергенции, а не дивергенции, и, следовательно, правильнее считать эти подотряды отрядами. В подтверждение своей гипотезы авторы приводят многочисленные морфологические различия между усатыми и зубатыми китами. Проблема дифилитического происхождения китообразных разработана еще недостаточно и требует дальнейшего изучения. Наиболее древние остатки усатых китов известны из верхнего олигоцена, а зубатых — из значительно более древних отложений — верхнего эоцена. Предки усатых китов имели зубы, о чем свидетельствуют не только палеонтологические, но и эмбриологические данные. Систематика китообразных к настоящему времени изучена недостаточно полно. Как уже сказано выше, даже целостность всего отряда ставится некоторыми учеными под сомнение. Объемы некоторых семейств также не установлены окончательно. В отряде китообразных обычно различают 2 современных подотряда: зубатых китов — Odontoceti, Flower, i867; усатых китов — Mysticeti Flower, 1864. В фауне СССР представлены оба подотряда. 1. В отряде можно выделить китообразных малых размеров с длиной тела 1,1- 12 м., средних - с длиной тела 12,1-22 я крупных —22.1—§3 м. 2. По-видимому, фонтан может образовываться и в результате конденсирования пара в теплом выдыхаемом воздухе (в высоких широтах), и в результате захватывания и распыления воды, которая остается при выныривании кита в ложбине, где располагается дыхало. | |
| Категория: Подотряд зубатых китов | (06.09.2013) | |
| Просмотров: 1044 |
